Jonathan Osborne and Dr. Michael Whiting, Integrative Biology
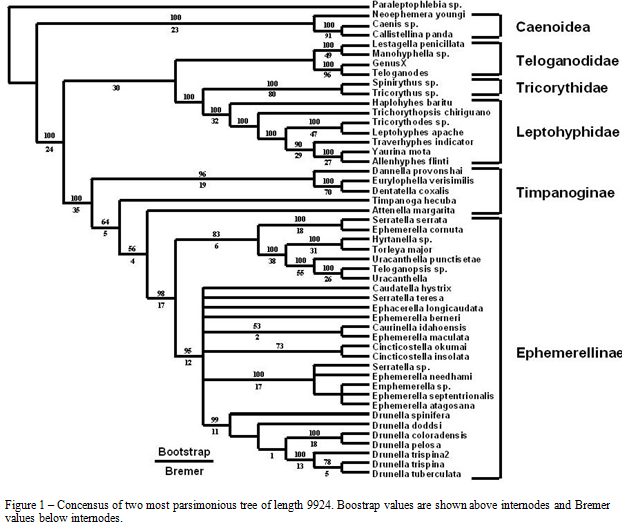
A prior analysis of the families of Ephemerelloidea, based on 49 morphological characters, supported Ephemerellidae diverging basal to all other Ephemerelloid families (McCafferty and Wang, 2000). The family Ephemerellidae contains twenty genera in two subfamilies (McCafferty and Wang, 2000) and comprises the largest family among the pannote mayflies. Members of this family are distributed throughout Holarctic and Oriental regions. The presence of certain species of the family Ephemerellidae in streams has been used to indicate high water quality. For example Drunella doddsi is shown to have a low tolerance to different environmental conditions such as high levels of alkalinity and sulphates (Mangum and Winget, 1991). Previously, members of the five genera currently included in the subfamily Timpanoginae were placed as subgenera of Ephemerella. McCafferty and Wang (1994) referred to this group of taxa as the Timpanoga complex and were the first to raise a number of the included subgenera to generic status and further differentiated them from other members of Ephemerellinae. Their conclusions were based primarily on the absence of lamellate gills on metasomal segment 3 in larvae and the absence of corresponding vestigial gill sockets in subimagos and adults. The Timpanoga complex was further modified resulting in the subfamily Timpanoginae, which includes three tribes and five genera (McCafferty and Wang, 2000; McCafferty, 2000). This study will investigate the relationships previously discussed here, look at higher level relationships within Pannota, and test the monophyly of some important Ephemerellid genera.
Taxa sampling included 6 of the 10 currently recognized pannote mayfly families as well as members from16 of the 18 extant Ephemerellid genera. Multiple species were sampled from Drunella, Ephemerella, Cincticostella, Serratella, and Uracanthella for testing of monoplyly. In all, sampling included 6 families, 32 genera, and 48 species.
The phylogeny was constructed based on 124 morphological characters and DNA sequence data from the following genes: 18S rDNA, 28S rDNA, 12S rDNA, 16S rDNA, and Histone3. Insect tissue was preserved in 100% ethanol. After extraction with Qiagen DNeasy protocol, DNA was amplified via Polymerase Chain Reaction (PCR), with oligonucleotide primers designed to amplify specific gene regions. Target product was verified via agarose gel electrophoreses and sequenced on an ABI 3100 automated DNA Sequencer. Correction of nucleotide sequences and assembly of contiguous sequences were performed using the computer program Sequencher 4.1. The data were analyzed via direct optimization under parsimony as implemented in POY (Gladstein and Wheeler, 1999). The gap/tranversion/transition ratio was set to unity (1:1:1). Bootstrap values were calculated from the implied alignment in PAUP* 4.0b4a (Swofford, 2002). Bremer values wer calculated using TreeRot and PAUP* 4.0b4a.
Analyses resulted in 2 equally parsimonious trees. Figure 1 is a strict consensus of these trees. The superfamilies Caenoidea and Ephemerelloidae were supported as monophyletic with high Bootstrap and Bremer values. Ephemerellidae is shown diverging basally to the remaining Ephemerelloidea as predicted by McCafferty and Wang (2000). The families Tricorythidae, Teloganodidae, and Leptohyphidae were recovered as monophyletic. Ephemerellidae is monophyletic. Timpanoginae is shown as paraphyletic with low to moderate support values. The subfamily Ephemerellinae was recovered, but many of the genus level relationships within this subfamiliy were difficult to determine. This same problem occurred in preliminary anaylises, but with the addition of more data the relationships became better resolved. Of the genera tested for monophyly within Ephemerellinae, Drunella and Cincticostella were supported as monophyletic. Results showed Serratella as a polyphyletic group spread throughout the Ephemerellinae. Uracanthella formed a paraphyletic group with the inclusion of an undetermined species of Telopanopsis. Ephemerella, as expected, is also polyphyletic; although, one clade including E. septentrionalis, E. needhami, E. atagosana, and one undetermined Ephemerella species did form a well supported clade with the inclusion of an undetermined Serratella species.