Sarah Walker and Dr. C. Riley Nelson, Department of Biology
Many beta diversity indicies, like most analytical tools in ecology, abstract the known relative abundances and species presence into outputs primarily useful for obtaining p-values and little else. Therefore, the purposes of this proposed beta diversity method were to simplify currently used similarity analyses for visualization and quantification of beta diversity, investigate its use in seasonal succession and other gradients in communities, and demonstrate an application of these methods in terms of both taxonomic and functional richness.
Materials and Methods
From 2003 to 2007, the American Fork River in central Utah was sampled for terrestrial adult forms of aquatic macroinvertebrates. With emphasis on the 2004 component of this data set, I developed a simple method to assess time or place of community turnover along a temporal or spatial gradient using similarity indicies. Initial development of these ideas was inspired by communication with Dr. John Longino of Evergreen State College.
Construction of so-called Sierra plots is fairly straightforward in that a matrix is composed giving the Morisita-Horn or Chao-Sorenson similarity values for each site as it relates to each other site, including each site’s similarity to itself which is 100% or a value of 1. From this matrix, a line graph is composed treating each site (or date) as a line or data series, thereby charting its similarity to each other site as well as the rate of turnover between each site or sample. The slope of the similarity lines composing Sierra plots are important for
determining rates of community turnover, or beta diversity. Slopes were calculated for each segment of the line to assess rates of change between sites or dates.
In order to further assess similarities between sites and samples, Non-Metric Multidimensional Scaling (NMDS), Similarity Percentages (Simper), and Analysis of Similarities (ANOSIM) analyses were conducted using Primer 6. The Bray-Curtis similarity index was selected for use in comparisons. Factors including month the samples were taken in, proscribed plateau community, which site the sample was collected at, and dominant character states for functional traits were entered as variables to assess influences on similarity. To assess changes in functional trait structure of the community, two-way nested ANOSIM analyses were performed with the specific trait in question nested within either site or month classification. ANOSIM outputs were used as additional analyses because this method is currently common in the literature and would therefore provide a comparison to Sierra plot findings.
Results
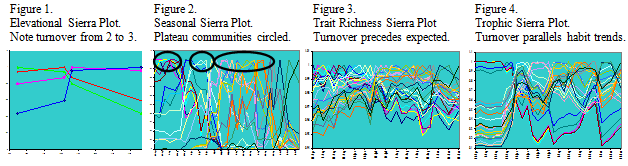
Beta diversity measures using Sierra plot slopes are indicative of community
turnover rates between sites. The sharpest rate of change occurs between Site 2 and 3
(Fig. 1) which are, counterintuitively, closest in both elevation (44 m difference) and
distance (2.49 km difference). The transition between Sites 2 and 3 is 9.686 to 13.554 times greater than change between other sites. Results were similar when using the Morisita-Horn index and either species or family groupings . Functional traits, particularly voltinism and habit type, and trait richness also followed this trend of sharp turnover between sites 2 and 3.
Seasonal community changes, also visualized by Sierra plots (Fig. 2), show three prominent communities that consistently occur over the course of each year and at each site. Some variation occurs in phenology of the plateaus, but most are centered around dates in April (“Winter”), June (“Spring), and August (“Summer”). Plateau communities were also supported by clustering analysis performed using Primer 6.
Functional traits, as measured by the dominant character state of a sample, differed across time with little correlation to turnovers in species or family richness. Trait richness, as examined by treating each functional trait as if it were a species, showed one community turnover between June and July (Fig. 3). This preceded a turnover of the same samples in taxonomic richness, both family and species, which occurred in July. These communities delineated by taxonomy also experienced an earlier turnover in June which the functionally defined community did not.
Individual traits exhibited different seasonal patterns. Both trophic groups and habit types showed a strong turnover between May and July (Fig. 4). However, life cycle strategy and development rate both had a strong turnover in mid-June. Interestingly, voltinism was highly variable from April to May but then had a strong plateau community thereafter. Other traits such as size and drift mobility also had interesting turnover patterns.
Discussion
The appearance of what I have termed ‘plateau communities’ was an unexpected result of a large study intended to survey the aquatic insect community of the American Fork River in Utah. The formation of a cohesive seasonal community opens questions about community structuring. Why, for instance, do certain species occur together and in what ways to they overlap functionally? Additionally, how do the species within a plateau community relate ecologically and evoluationarily? And do different plateau communities exhibit similar functional structure and do what degree?
The discipline of ecology, currently dominated by NMDS plots and clustering analyses, can benefit from investigating new and simpler approaches to analyze community turnover that are more accessible to students, researchers, and conservationists alike.
Reference
- Longino, J. T. 2006. Richness and faunal turnover of ants (Hymenoptera, Formicidae) on the Barva Transect. Entomological Society of America Meetings. Indianapolis, IN.; Longino, J. T. 2007. Personal correspondence (email). October 24, 2007.