Jennifer Rasmussen and Dr. Keith Crandall, Biology Department Chair
Main Text
Shrimp-like decapods–including sergestoids, penaeoids, carideans, procaridids, and stenopodideans–are a species-rich and morphologically diverse group of crustaceans. Some taxa, such as the penaeoids and carideans, are economically important, bringing in billions of dollars each year to world fisheries. The economic importance of this group, together with their distinctive morphology and ecological diversity, makes shrimp-like decapods popular research subjects in all fields of biology.
Understanding the phylogenetic relationships among shrimp-like decapods can play a significant role in conservation efforts and biodiversity estimates. This can be particularly important for commercially important or habitat-threatened groups. Penaeoid shrimp are monetarily important and heavily harvested for human consumption. Procaridid shrimp, recently discovered in 1972, can only survive in anchialine pools. These rare and unique habitats often contain endemic taxa threatened by invasive species and habitat loss.
Many researchers have investigated the phylogenetic relationships among decapod crustaceans using morphological and molecular data, however these studies suggest conflicting hypotheses of evolution and lack a robust sampling of shrimp-like taxa (Ahyong & O’ Meally, 2004; H. Bracken et al., in press; Dixon, Ahyong, & Schram, 2003; Porter, Perez-Losada, & Crandall, 2005; Scholtz & Richter, 1995; Toon, Finley, Staples, & Crandall, 2009; Tsang, Ma, Ahyong, Chan, & Chu, 2008). Although the superfamily Penaeoidea and infraorders Caridea and Stenopodidea are frequently present in these analyses, Sergestoidea and Procarididea are rarely included, possibly due to the inaccessibility of specimens.
This project intended to use nuclear protein-coding genes (PEPCK, NAK) to resolve inner-node relationships among the major shrimp-like decapod lineages including Sergestoidea, Caridea, Penaeoidea, Stenopodidea and Procarididea. We planned to use these genes to support a relationship between the pleocyemate taxa Stenopodidea, Procarididea, and Caridea. We wanted to highlight monophyletic and polyphyletic assemblages, while also examining congruence or incongruence between past and present hypotheses of evolution in shrimp-like decapods. PEPCK and NAK, protein-coding genes were intended to solve inner-node discrepancies, however they have yet to be resolved. However, here we present the first molecular phylogeny for shrimp-like decapods, including all major lineages. We combined nuclear and mitochondrial sequences, to investigate the relationships among 3 pleocyemate infraorders, 2 dendrobranchiate superfamilies, 35 families, 81 genera, and 107 species.
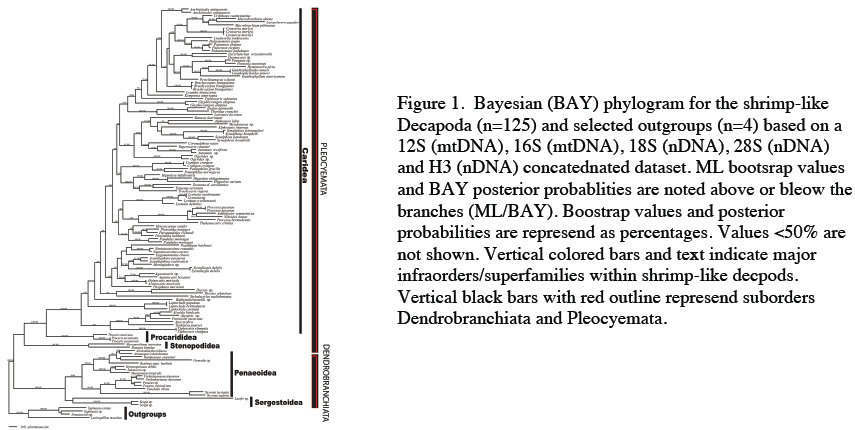
In total we included 122 16S, 99 12S, 129 18S, 103 28S, and 117 H3 sequences. Results strongly support the monophyly of the suborder Dendrobranchiata (ML bootstrap support (bs) = 100%, pP = 1.0), however there is little support for the superfamilies within this group (Penaeoidea (bs = 50%, pP = 0.84); Sergestoidea (bs = < 50%, pP = < 0.50; Fig. 1). Within the Dendrobranchiata, the genus Lucifer grouped within the Sergestoidea (Fig. 1) or as a basal-root to the remaining dendrobranchiate taxa (not shown).
Although findings do not statistically support a relationship between the pleocyemate taxa Stenopodidea, Procarididea, and Caridea, the Procarididea were confirmed as the sister taxa to the Caridea (bs = 100%, pP = 1.0; Fig. 1). In alternative ML and BAY topologies, the stenopodideans were basally positioned to the Dendrobranchiata. There is statistical support for the following pleocyemate infraorders: Caridea (bs = 100%, pP = 1.0), Procarididea (bs = 100%, pP = 1.0), and Stenopodidea (bs = 100%, pP = 1.0).
Resolving infraordinal relationships continues to be the most disputable and unsettled issue in decapod evolution. Although our study offers strong support for some infraordinal relationships, the phylogenetic position of other groups remains uncertain.
I would like a wide variety of friends and colleagues who have helped collect shrimp taxa from around the world over the many years. I would also like to extend a special thanks to Keith Crandall and Heather Bracken, for their support and knowledge during this project.
References
- Ahyong, S. & O’ Meally, D. (2004) Phylogeny of the Decapoda Reptantia: Resolution using three molecular loci and morphology. The Raffles Bulletin of Zoology, 52, 673-693.
- Bracken, H., Toon, A., Felder, D. L., Martin, J. W., Finley, M., Rasmussen, J., et al. (in press) The Decapod Tree of Life: compiling the data and moving toward a consensus of decapod evolution Arthropod Systematics and Phylogenetics.
- Dixon, C. J., Ahyong, S. & Schram, F. R. (2003) A new hypothesis of decapod phylogeny. Crustaceana, 76, 935-975.
- Porter, M. L., Perez-Losada, M. & Crandall, K. A. (2005) Model-based multi-locus estimation of decapod phylogeny and divergence times. Molecular Phylogenetics and Evolution, 37, 355-369.
- Scholtz, G. & Richter, S. (1995) Phylogenetic Systematics of the Reptantian Decapoda (Crustacea, Malacostraca). Zoological Journal of the Linnean Society, 113, 289-328.
- Toon, A., Finley, M., Staples, J. & Crandall, K. A. (2009) Decapod phylogenetics and molecular evolution. In: J. W. Martin, K. A. Crandall & D. L. Felder (Eds), Decapod Crustacean Phylogenetics (Crustacean Issues 18). CRC Press, Bocan Raton, FL, pp. 14-28.
- Tsang, L. M., Ma, K. Y., Ahyong, S. T., Chan, T. Y. & Chu, K. H. (2008) Phylogeny of Decapoda using two nuclear protein-coding genes: Origin and evolution of the Reptantia. Molecular Phylogenetics and Evolution, 48, 359-368.