
Andres Mallipudi with Erwin Barria, Carlos G. Jara, and Keith A. Crandall
Abstract
The biologically diverse region of Patagonia is under constant threat of urban development from both external and internal sources. Gaining a thorough understanding of the ecological roles of this area’s organisms is essential to preserving Patagonia’s rich biodiversity. Researching the differences between Southern Chilean crab subspecies led to a better understanding of this taxon, which will help determine its ecological role and will contribute to gaining a deeper understanding of the ecological environment of the area. Examining genetic relationships, divergence times, and morphometric variations between two different types of southern Chilean crabs (A. denticulata denticulata and A. denticulata lacustris) allows us to connect species diversification to ecological factors or geological events (i.e. Last Glacial Maximum) that occurred in the past. The information gathered throughout this project will help direct preservation efforts to higher priority areas by determining the biodiversity levels throughout the region (Perez-Losada et al.) This knowledge will aid in curtailing the harmful effects that development will cause by furthering our understanding of the function of these organisms within their ecosystem.
Introduction
Freshwater crabs of the family Aeglideae are restricted to Southern South America and are present in Chile, Brazil, Bolivia, Uruguay, Paraguay and Argentina. The entire family is a part of the genus Aegla, of which there are 63 known species, 36.5 percent of these organisms are recognized as being under threat or endangered by the IUCN Red List Criteria. There are 5-10 additional species which have been observed but are not yet classified (Bond-Buckup). The incredible variance among this single genus showcases the vast diversity within the region of Patagonia.
There is currently economic pressure, both domestic and international, to develop this region; its broad diversity levels that are unique to thi virtually untouched landscape are under constant threat of disturbance. Our research has led to a better overall understanding of the biodiversity in this region which will help minimize the effect that future development will have on the area. Furthering our knowledge of the organisms in this area will allow us to better manage and reduce the harmful effects that development will cause.
Materials and Methods
The research took place in and around Valdivia (39° 48′ 0″ S, 73° 14′ 0″ W), which is located in southern Chile in the biologically diverse region of Patagonia. Valdivia has several lake and river systems which were created about 8,000 years ago when the glaciers located in this region melted. Pleistocene glacial cycles have significantly impacted the distribution of freshwater species in Southern Chile (e.g., Xu et al. 2009). The freshwater crab species Aegla denticulata denticulata can be found in the river systems formed from melting ice sheets (Nicolet 1849), and in 1989 a subspecies was found in the connecting lake systems (Jara, pers. comm.).
We took 328 samples from multiple locations. The locales in which we conducted our field work were Arroyo Chesque, Arroyo Puquitre, Arroyo Quilguen, Arroyo Forrahue, Rio Collilelfu and Lago Rupanco. Minimal previous work has been done showing some molecular and physical differences between these subspecies, but our research focused on the data gathered through genetic analysis and geometric morphometric techniques.
We will analyze how genetic variation differs between the lake and river subspecies by examining molecular divergence on three genetic loci (EF1 intron, 16S, COI). Total genomic DNA will be extracted from the organisms’ muscle tissue and the targeted gene regions will be amplified by means of the polymerase chain reaction (PCR). The computer program Sequencher (GeneCodes, Ann Arbor, MI, USA) will be used to assemble, edit and clean the desired sequences. The relative timing of divergence is estimated using Bayesian molecular dating methods. Using a strict molecular clock, which will be calibrated by a predetermined substitution rate, we will be able to determine the rate of change among the branches.
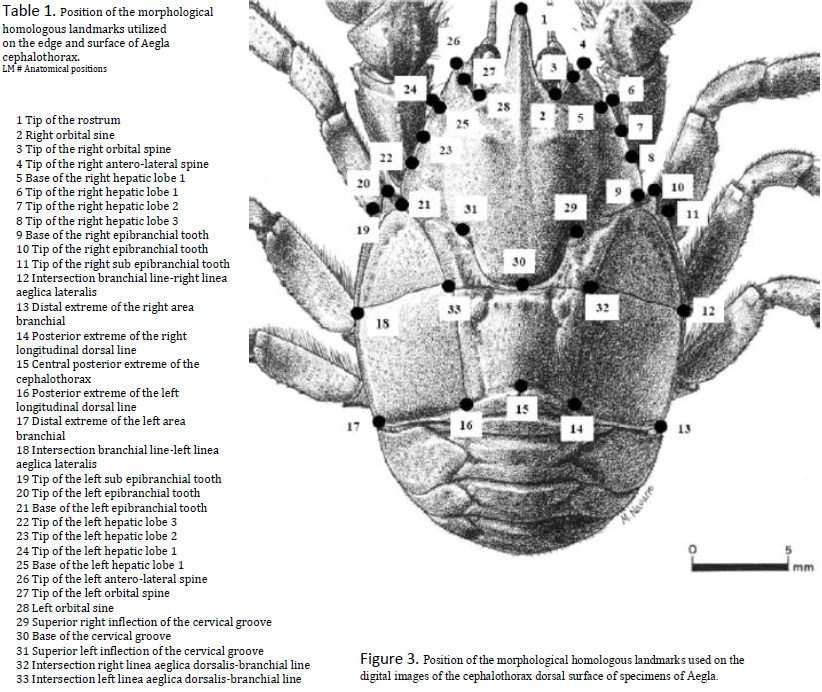
The use of geometric morphometrics is a method which quantifies the observational differences that exist between these two crab subspecies (Zelditch, et al, 2004). This allows researchers to determine whether or not the measured physical differences between the organisms are statistically significant, and not simply the result of pure chance. Through the process of morphometrics we chose several similar landmarks that exist in both subspecies and measured the distance between these landmarks. This distance provides the numerical values for each crab that provides the data necessary to demonstrate the differences that exist between the two subspecies.
Examining the genetic relationships and divergence times between these two closely related subspecies allows us to connect species diversification to ecological factors or geological events (i.e. Last Glacial Maximum) that occurred in the past. Significant phylogenetic differences, determined through Bayesian analysis, demonstrate that this area is a high priority target for conservation efforts. Divergence times close to estimates of when deglaciation occurred suggest that speciation may have happened during this time. Morphological differences, such as larger overall size and longer rostrums in A. denticulata lacustris, would indicate that the species adapted to its particular environment. This combination of genetic data and morphometric analysis allows us to tie the differences in genetic diversity and morphology to the differences in habitat.
Results
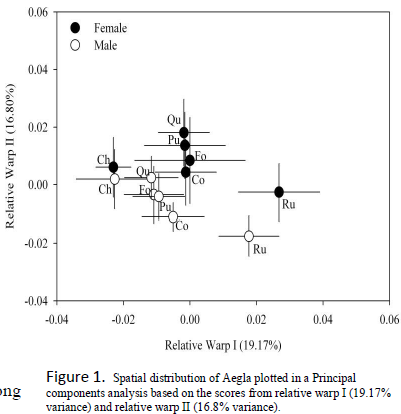
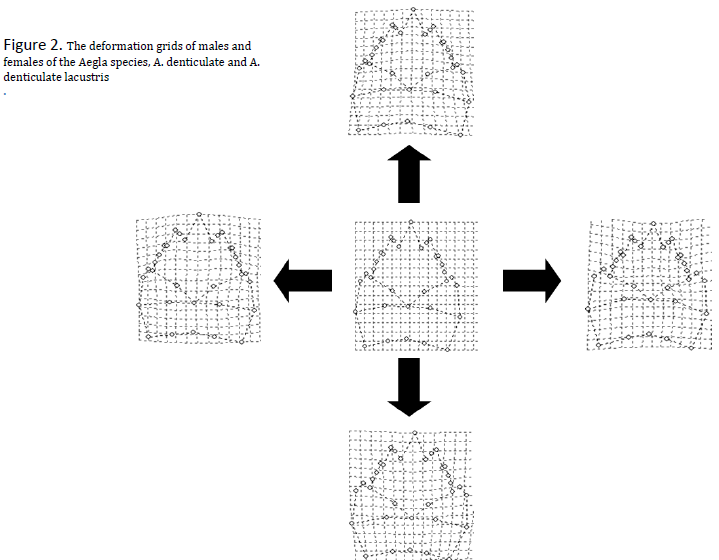
The measurements along the x-axis (Relative Warp I, from here on knows as RWI) were an analysis of the configuration of landmarks on the posterior half of the cephalothoracic border, while those along the y-axis (RWII) took into account variations along the rostral spine (see figure 3 for additional clarification). As RWII values increase, position along the y-axis increases. Likewise, a high RWI value would correspond to a more rightward position on the corresponding graph (Figure 1). The total variance of 35.97% is explained by the first two relative warps (RWI: 19.17%, RWII: 16.80%). The two-way analysis of covariance (ANCOVA) used to compare relative warps (Figure 2), reported statistically significant differences when using sex and population as sources of variation. Measurement of the centroid was used as a control. The a posteriori test applied to our hypothesis test revealed that four groups appeared in regards to RWI values when considering statistically significant (p<0.05) differences in morphological variation. The population with the smallest RWI value, Arroyo Chesque (Ch), form one group. The populations from Arroyo Quilguen (Qu), Arroyo Forrahue (Fo), Arroyo Puquitre (Pu) and Rio Collilelfu (Co) comprise another group. The groups with the largest RWI values, the female and male populations from Lago Rupanco, make up the last two intrinsically similar but statistically distinct groups. With the RWII analysis, the female population differed significantly from the male population in all cases except for the group from Arroyo Chesque. The ordering for RWII analysis was a function of sex. The groups in the RWI analysis also formed as a result of sexual dimorphisms, but were organized primarily as function of population.
Discussion
The statistical variance demonstrated by the relative warp analysis in this study (35.97% is low relative to values reported (88.97% and 62.97%, respectively) in previous studies that analyzed inter-specific morphological variation (Erwin Barria 2008, Giri & Collins 2004). These previous reports were more heavily weighted towards inter-specific differences in comparison to this study. In addition, the results indicate that the variables that predict morphological variation in the comparative experimental units for this study are more weighted towards sexual dimorphisms. As was evident with the RWI analysis, the inter-population differences were unable to explain the variation among the corporal cephalothorax plane. These results can be partially explained by the segregation of the Lago Rupanco population from the other groups, which accounts for a large portion of the variance. The morphological variations of the Lago Rupanco populations compared to the other groups are evident and occurred as a result of interspecific comparison.
The analysis of morphological variation between species is complete, but the molecular analysis has not yet commenced. Once these data are acquired, we will test the hypotheses on how convergent evolution (distinct and unrelated lineages developing similar traits) and speciation (process through which new species arise) affected these subspecies differently (Tollrian et al 1998). Does the lacustrine species differ significantly from the riverine population? Are the differences between crabs across different lakes statistically significant? Do similarities exist between the crabs in nearby lake and river systems? Evidence showing similarities between lake crabs (as opposed to similarities existing between crabs in interconnected lake and river systems) would indicate that convergent evolution has taken place. If the latter results are acquired we may assume that either not enough time has passed in order for convergent evolution to take place, or that each river-lake system will develop into its own subspecies and convergent evolution will not occur.
Acknowledgements
We would like to thank Nicole Wood and Dr. Heather Bracken-Grissom for their help with project coordination, lab work, and all of their input and advice on this draft. This work was made possible through grants from Brigham Young University’s Office of Research and Creativity and through the Department of Biology.
Scholarly Sources
- Barria, Erwin M., Roger D. Sepúlveda, and Carlos G. Jara. ―Morphologic Variation in Aegla Leach (Decapoda: Reptantia: Aeglidae) from Central-Southern Chile: Interspecific Differences, Sexual Dimorphism, and Spatial Segregation.‖ Journal of Crustacean Biology, 31(2):231-239. 2011.
- Bond-Buckup, Georgina, Carlos G. Jara, M. Perez-Losada and Keith A. Crandall. ―An assessment of global diversity of the freshwater crab Aeglidae (Decapoda: Anomura).‖ Hydrobiologica 595 (2008), 267–273
- Jara, Carlos G. ―Aegla-Denticulata-Lacustris new-subspecies from Lake Rupanco Chile crustacea decapoda anomura aeglidae.‖ Biological Society of Washington. Volume 102. Issue 2 (1989): pages 385-393.
- Giri, Federico and Collins, Pablo Agustin(2004) ‘A geometric morphometric analysis of two sympatric species of the family Aeglidae (Crustacea, Decapoda, Anomura) from the La Plata basin’, Italian Journal of Zoology, 71:1, 85 — 88.
- Perez-Losada, Marcos, Georgina Bond-Buckup, Carlos G. Jara, K. A. Crandall 2009. ―Conservation assessment of southern South America freshwater ecoregions on the basis of the distribution and genetic diversity of crabs from the genus Aegla.‖ Conservation Biology 23(3), 692–702.
- Tollrian, Ralph, and C. Drew Harvell. The Ecology and Evolution of Inducible Defenses. New Jersey : Princeton, 1998.
- Xu, J.W., Pérez-Losada, M., Jara, C.G. & Crandall, K.A. 2009. Pleistocene glaciation leaves deep signature on the freshwater crab Aegla alacalufi in Chilean Patagonia. Molecular Ecology. 18: 904-918.
- Zelditch, Miriam Leah, Donald L. Swiderski, H. David Sheets, and William L. Fink. Geomeric Morphometrics for Biologists: A Primer. New York; Maple- Vail, Kirkwood, 2004.