Heather Finch and Dr. Phil Allen, Department of Plant and Wildlife Sciences
Invasive species such as Bromus tectorum, commonly known as cheatgrass, occupy over 40 million hectares of land within the interior portion of the United States. The naturally occurring pathogen Pyrenophora semeniperda has been discovered to kill the seeds of this invasive species. We believe that it could be used as a potential biocontrol within the field. But like many pathogens, it has a catch. P. semeniperda preferentially kills dormant seeds.
The explanation to the pathogen’s preference is best explained in a paper published in 2007. In this paper, the answer as to why dormant seeds are preferentially killed is explained by “The Race for Survival Hypothesis”. This hypothesis states that germination rate is a critical factor that determines whether or not seed death will occur. In other words, seeds that germinate quickly escape the pathogen. If the seeds are slow germinating (for example, dormant seeds), the pathogen has the ability to beat germination and kill the seed. In contrast, fast germinating seeds such as non-dormant seeds would “outrace” the pathogen because they would germinate too quickly. In the field, non-dormant seeds germinate when they have enough water after a summer rain storm, but does it always rain enough for seeds to germinate? If they didn’t obtain enough water, would the pathogen be able to “win the race” and kill the seed before it had a chance to germinate? About a year ago, I obtained data that suggested that indeed, P. semeniperda can infect and kill non-dormant seeds in conditions where limited water is present.
To further study how this occurs, I conducted an independent experiment using additional variables than those that I used in my first independent study. To determine if the pathogen had a wider range of temperatures in which it could kill seeds, I added four additional temperatures to incubate seeds at. I also varied the time seeds were incubated in low water potentials (varying conditions of water available to the seed) in order to determine how much time was necessary for the pathogen to kill the seeds. I hypothesized that pathogen- inoculated seeds incubated for shorter periods of time in a low water potential treatment combined with colder temperatures would result in higher seed mortality.
To disprove this hypothesis, I conducted a factorial experiment using both dormant and non-dormant, inoculated and un-inoculated seeds, three water potential levels (0 MPa, -1.5 MPa, -2.0 MPa), five temperatures (5⁰, 10⁰, 15⁰, 20⁰, 25⁰C.) and four different time periods (7, 14, 21, 28 days). I used 2 reps of 50 seeds, allowing for there to be 480 dishes, or 24,000 seeds total. For each time period, (120 Petri dishes containing 50 seeds each in each period) I incubated seeds in low water potentials using a chemical called Polyethylene Glycol. When mixed with water, this chemical inhibited seeds from germinating. The more negative the level, the more difficult it was for seeds to germinate. On days 2, 4, 7, 11, 14, 21, and 28 I recorded whether the seed exhibited disease (as indicated by the presence of black protrusions, or stromata, from the endosperm of the seed) or had germinated. After the periods of 7, 14, 21, or 28 days had passed, I transferred all seeds to dishes containing pure water and incubated them for an additional 28 days.
While obtaining data, it was noticed that a contamination was present in some of my dishes. We later discovered that a Fusarium organism was infecting my seeds and making data unobtainable at 25 degrees C. Even though I experienced some difficulties with this contamination, I was still able to obtain valid data from the other temperatures.
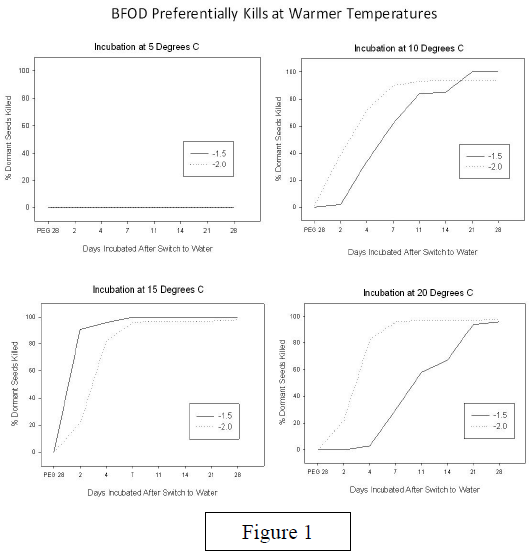
In conclusion, my hypothesis that pathogen- inoculated seeds incubated for shorter periods of time in a low water potential treatment combined with colder temperatures proved to be incorrect. In fact, inoculated seeds that are incubated in cold temperatures such as 5 degrees C. do not exhibit disease at all. It seems that the pathogen does not work best in colder temperatures such as 5 and 10 degrees C., suggesting that this pathogen is programmed to best work in the Fall (figure 1). The cold temperatures actually seemed to encourage dormant seeds to germinate. In addition to temperature, I was able to determine that 14 days incubated at low water potentials was sufficient for over 50% of non-dormant seeds to be killed (figure 2). Any additional incubation time at a low water potential level gave advantage to the pathogen. Shorter time periods were not a sufficient amount of time for significant numbers of non-dormant seeds to be killed.
These results indicate that early season rain storms inadequate for germination can allow the pathogen to “win the race” as long as there is adequate time for seeds to be left at low water potentials in the field. Temperatures such as 15 and 20 degrees C. are preferred by the pathogen in order to obtain highest mortality of both dormant and non-dormant seeds.
After obtaining this data, I was able to attend the Ecological Society of America conference in Austin, Texas to present my results. We are currently working on a paper in order to publish my results. I am also planning on obtaining a Masters degree, using this data as part of my research to further understand how Pyrenophora Semeniperda can kill non-dormant seeds.