Chris Cowdell, Departments of Food Science Nutrition/Molecular Biology
Object
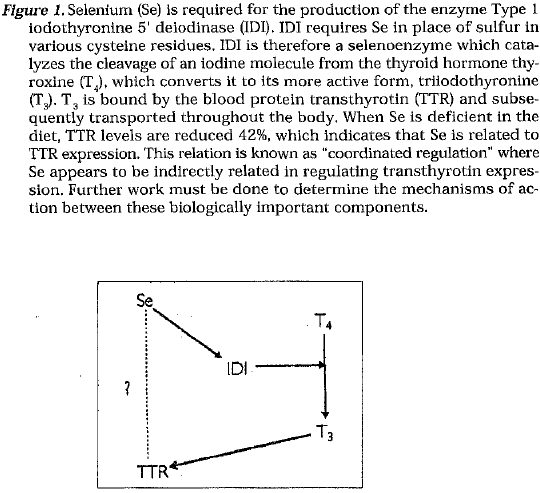
Various genes have been discovered which are regulated by the dietary Intake of the trace element Selenium (Se).Some of these genes code for such enzymes as glutathione peroxidase (GP J Type I iodothyronine 5′ delodinase (IDI), and selenoproteln P (Se-P). GP, is an enzyme (a selenoenzyme) which Incorporates Se in place of sulfur within residues of the amino acid cysteine. GP, Is active In the assistance of free radical elimination throughout the body by converting hydrogen peroxide (H,02 ) to oxygen (02) and water (H20). IDI is a selenoenzyme which catalyzes the conversion of the thyroid hormone thyroxine (T 4) to its more active form triiodothyronine (TJ. Se-P Is a plasma selenoenzyme whose function has not yet been elucidated.
The sole focus of this research project is to identify a gene in rat liver that is differentially expressed due to the amount of Se In the diet. This gene may be an uncharacterized gene or a previously characterized gene which was not previously known to be regulated by dietary Se intake.
Methods of Research
Rats and Diet: Two populations of weanling male Sprague-Dawley ats were used. One group was fed a normal diet and the other fed a elenium (Se)-deficient diet for approximately 90 days. The rats were kept in steel cages with constant access to both food and water. Both diets consisted of a torula yeast-based meal using corn oil and sucrose to provide lipid and carbohydrate. The individual vitamins and minerals necessary for rats were added separately. The Se-adequate diet was supplemented with 0.5 ppm Seas Na,Se03, and the Se-deficient diet had no added Se except for the minute fraction found in the torula yeast.
RNA Isolation: Total RNA was isolated from rat livers. The rats were decapitated, after which the liver was Immediately excised, frozen in liquid nitrogen and stored at -SOT until the RNA was isolated. The isolation was performed using the commercially available reagent STAT -60, which was based on the method of Chomzynskl and Sacchi. The RNA was then treated with RQI RNase-free DNase (Promega) to remove any DNA that could Interfere with the subsequent reactions. The RNA was then stored at -80’C.
Method of Identifying Differentially-Expressed Genes: To identify differentially- expressed genes, a modified version of differential display was used, a technique originally devised by Liang and Pardee. MMLV reverse transcrlptase (Gibco BRL!Life Technologies) was used to reverse transcribe Seadequate and Se-deficient RNA, using oligo-dT (Perkin Elmer) as a primer. The oligo-dT was then removed via Microcon-30 microcon-centrators (Amicon). This step was required because oligo-dT would act as a primer In the subsequent polymerase chain reaction (PCR) steps, producing smears within gels instead of the desired discrete bands.
PCR was then used to selectively amplify the reverse transcription products. I 0-mers (Operon Technologies, Inc.) were used as primers. The Stoffel fragment of Taq DNA polymerase (Perkin Elmer) were used to catalyze the PCR reactions. 35S-labeled dATP was added to the reactions to radiolabel the products. The PCR reaction cycled 40 times (94’C for 30 seconds, 40′ C for 1 minute, and 72′ C for 2 minutes).
The PCR reactions were then electrophoresed through a denaturing polyacrylamide sequencing gel (6% polyacrylamide, 6M urea). Lanes of Se-adequate and Se-deficient reactions were run next to one another for each primer pair used. After exposure to and the development of the autoradiography film, the film was analyzed for differences in band intensity (indication of differentially-expressed genes). A band was present in both adequate lanes. It was not present in the deficient lanes, which indicated that a gene was being expressed with adequate dietary Se, but not with deficient levels of dietary Se.
Isolation of Specific DNA/Verification of Differential Expression: The areas of the acrylamide gel which corresponded to the differentially expressed genes were then cut out. The DNA was then isolated from the gel slices and reamplified by PCR using the same I 0-mers and enzymes used in the initial PCR reaction. The amplified DNA was then radiolabeled with 32P-dCTP using the Random Primed DNA Labeling Kit (Boehringer Manheim). The labeled DNA was then used to probe a Northern blot. The same Se-adequate and Se-deficient RNA used in the reverse transcription reactions were analyzed. G3PDH was used as a control probe.
One problem encountered with the use of differential display and its derivatives is the presence of comigrating species. Many times a single band from a differential display gel contains more than one species of DNA which appears to be of the same length. Comigrating species are manifest when more than one band is present on a Northern blot. Although the species of DNA are the same length, the RNA with which they hybridize are of different lengths.
Comigrating species were manifest, producing the need for a procedure called ‘affinity capture’ to isolate the species which was differentially expressed. The area of the northern membrane which corresponded to the probe which was previously used was excised. The DNA was stripped off this small piece of membrane and reamplified via PCR.
Cloning the Differentially-Expressed DNA: After the DNA was isolated, it was cloned using theTA Cloning Kit (Invitrogen), which uses the pCRII vector. A plasmid mini prep was then performed using the Qiaprep Spin Plasmid Kit (Quiagen). The insert in the plasmid was isolated via restriction digestion with the appropriate restriction endonucleases. The plasmid insert was then radiolabeled with 32P-dCTP and used to probe a Northern blot, which was necessary to verify that the DNA fragment in the plasmid was the differentially-expressed DNA which was previously isolated. G3PDH was again used as the control probe.
Sequencing and Screening: The insert in the plasmid was sequenced (via sequencing gel) and compared to known DNA sequences in GenBank at the National Libraries of Medicine. The differentially expressed gene corresponded to citrate transport protein (CTP) and transthyrotin (TTR), both found in rat liver. The full-length cDNAs for each of these genes were then obtained from the researchers who first isolated them. These cDNAs were then used to probe another northern blot, which verified that the fragment of DNA which was isolated from the Se-adequate rat livers was actually the same as the full-length cDNAs (CTP and TTR) obtained from the previously mentioned researchers.
Results
Preliminary quantitation showed that CTP was reduced by approximately 80-85% in the Se-deficient rat livers. These results are significant because CTP is a common, yet very significant protein required for proton transport across the inner mitochondrial membrane required by the body to sustain life. Because research in this area is relatively young, more work needs to be conducted in this area.
TTR is a protein previously known as pre-albumin. This protein is the major transport protein of thyroid hormone and (vitamin A) in the blood. Results show that TTR was reduced by 42% in Se-deficient rat liver compared to Se-adequate liver levels. This proves significant because a complete reduction of TTR would result in death.
The object of this research project consisted of identifying a gene regulated by dietary Se intake within rat liver. The project resulted in the identification of two genes which appear to be regulated by dietary Se. The focus of future work should concentrate on discovering the mechanisms of action for the regulation of Se on CTP and TTR levels.