Seth Taylor and Dr. Scott Steffensen, Psychology
Cocaine abuse is a serious societal problem. The 2006 Monitoring the Future survey of students showed that 2.5% of high school seniors had used cocaine in the past month, 5.7% in the last year, and 8.5% at least once in their lifetime. According to the 2005 National Survey on Drug Use and Health, 5.5 million Americans has used cocaine in the last year, twice as many as the number of people suffering from schizophrenia. Researchers have been working diligently to discover the means by which cocaine acts on the brain to cause such dependence. Treatments for addiction have improved as the knowledge of cocaine’s effects on the brain has increased, but much still remains to be discovered.
The current prevailing theory in cocaine addiction research involves the mesocorticolimbic dopamine (DA) system, which is responsible in the rewarding sensation for many natural behaviors (Koob et al., 1994). These behaviors include feeding (Ahn and Phillips, 2002) and drinking (Agmo et al., 1993). This system is also involved in the habit-forming actions of addictive drugs, including psychostimulants such as cocaine (Wise, 2004). It is believed that cocaine increases DA levels via blockage of the DA transporter, which removes dopamine from the extracellular space.
Cocaine also produces anesthesia by blocking sodium channels in neuronal membranes. These channels are needed to conduct signals between brain regions and from the brain to the rest of the body. It is possible that other systems are involved in the habit-forming actions of cocaine. Dr. Steffensen believes that one of these systems involves inhibitory γ-aminobutyric acid (GABA) producing neurons in the VTA. These neurons send inhibitory signals to many areas in the brain, including the areas involved in reward and memory. We hypothesized that cocaine reduces the ability of these neurons to inhibit other areas of the brain.
My main role was to conduct whole-cell patch clamp experiments to measure cocaine’s effects on these neurons. This is done by preparing very thin slices of rat brains (0.2-0.3 mm thick), and using fine tip pipettes, attaching an electrode to individual cells. Since neuronal cells transfer information through the conduction of electrical signals, this technique allows us to measure the activity of the cells by stimulating and recording the electrical responses of the cell. We wanted to determine the effects of various doses of cocaine on these inhibitory cells and the subsequent effect on the output cells of this brain region.
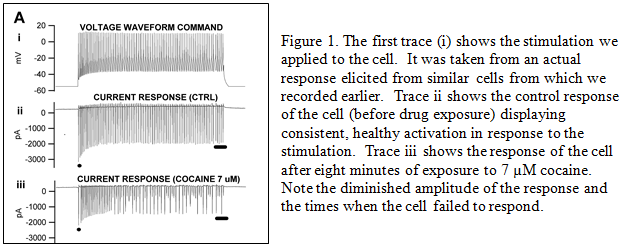
Results
We applied cocaine to these slice preparations at four different concentrations – 3, 7, 15, and 50 µM. We discovered that cocaine significantly reduced the sodium current in these neurons, which makes the neurons less active and less likely to inhibit the output cells.
Originally we planned to only run cocaine at three concentrations – 15, 25, and 50 µM. Our 15 µM results showed a significant effect and we therefore decided to decrease the concentration to lower levels to generate a more accurate dose-response curve. We were not able to achieve enough stable experiments at 25 µM to include them in my study. We also had originally planned to test the effects of methamphetamine, which shares some similar mechanisms of action with cocaine, but we determined that would not be necessary to show what we wanted to currently. This is an area that could be further researched to corroborate what we found.
These experiments implicated that cocaine does inhibit this subset of cells in the VTA. As these cells are thought to inhibit the dopamine-containing neurons, the cells most likely involved in mediating reward and value of behavior, we decided to test if cocaine led to a decrease in GABA input on the dopamine neurons. In a way, the GABA cells we tested first act as brakes on the activity of the dopamine neurons, and we believed cocaine was making the brakes less effective. This was not included in my original proposal, but fit in well with the purpose of the research.
We found that cocaine reduced GABA input to dopamine neurons. This effect was visible at all concentrations tested (7, 13, and 50 µM). This showed that the previously described effect of cocaine on GABA neurons led to a decrease in their influence on the major output cells of the VTA, the dopamine-containing cells involved in reward. These findings have been included, along with other research from our laboratory, in a revision of a paper we have submitted for publication in the European Journal of Neuroscience.
Sources
- Agmo A, Federman I, Navarro V, Padua M, Velazquez G (1993) Reward and reinforcement produced by drinking water: role of opioids and dopamine receptor subtypes. Pharmacol Biochem Behav 46:183-194.
- Ahn S, Phillips AG (2002) Modulation by central and basolateral amygdalar nuclei of dopaminergic correlates of feeding to satiety in the rat nucleus accumbens and medial prefrontal cortex. J Neurosci 22:10958-10965.
- Koob GF, Caine B, Markou A, Pulvirenti L, Weiss F (1994) Role for the mesocortical dopamine system in the motivating effects of cocaine. NIDA Res Monogr 145:1-18.
- Wise RA (2004) Dopamine, learning and motivation. Nat Rev Neurosci 5:483-494.