
Alexander Baxter and Dr, J. Dee Higley, Psychology
Introduction
Prenatally, androgens are responsible for masculinizing the developing male fetus. Prenatal androgen exposure (PAE) also shapes male brain functioning and organization, and is the main underlying influence underlying sexually dimorphism, to a certain extent even in adulthood. Through its effect on the brain, PAE can influence personality and behavior, likely contributing to gross sex differences in these traits, as well as within-sex variation (see Manning, 2011). One of the most commonly used biomarkers used to study PAE is the second-to-fourth-finger length ratio (2D:4D ratio). In humans, males tend to have a longer ring finger and shorter pointer finger (i.e., a 2D:4D ratio < 1), whereas females tend to have a longer pointer finger and shorter ring finger (i.e., a 2D:4D ratio ≥ 1). This sex difference is likely due to a higher concentration of androgen receptors in the ring finger relative to the pointer finger (Zheng & Cohn, 2010). Consequently, when testes produce androgens and masculinize the male fetus, the androgen receptors in the ring finger induce greater growth in the ring finger of males, compared to females. Research using 2D:4D research shows that in humans, low (i.e., male-typical) 2D:4D ratio is linked with variation in a variety of personality traits, including aggression, competitiveness, and temperament traits important to social dominance (see Manning, 2011).
Although many studies have investigated 2D:4D ratio in humans, relatively few studies have investigated 2D:4D ratio in nonhuman primates. Some studies suggest that the pattern of sex differences in 2D:4D ratio is reversed in some Old World monkey species (Abbott et al., 2012). Studies also show that in these species, 2D:4D ratio is associated with the social dominance that monkeys attain as adults (Nelson et al., 2010). Based on these studies, in a previous investigation (funded by a 2017 ORCA award), my mentor and I investigated 2D:4D ratio in the largest sample to date of rhesus monkeys, and found that these monkeys exhibit a sex difference in 2D:4D ratio, with males exhibiting higher 2D:4D ratio than females, a pattern that is reversed relative to the human pattern of sex differences in 2D:4D ratio (Baxter et al., in press). However, we did not have enough data on the alpha- and beta-ranking monkeys to explore the association between social dominance rank and 2D:4D ratio. Hence, with my project this year, I continued investigating 2D:4D ratio in rhesus monkeys, particularly in the alpha- and beta-ranking monkeys. I hypothesized that the alpha and beta subjects would exhibit higher (i.e., more male-typical for a rhesus monkey) 2D:4D ratio than the other subjects in their social group.
Methods
Monkeys’ fingers were measured while they were sedated as part of bi-annual health inspections. Following the procedures of Abbott et al., (2012) and Manning (2011), we measured monkeys’ 2nd and 4th digits along the midline of the finger, from the baseline crease of the finger most proximate to the palm to the tip of the finger. Measurements were taken at least twice, until two measurements were obtained within ±1.5 mm. The two closest measurements were used to determine average finger length. Subjects’ left- hand 2D:4D ratio and right-hand 2D:4D ratio were determined by the dividing the average length of their 2nd finger by the average length of their 4th finger. To date, we have measured more than 1,000 monkeys’ fingers, including 33 alpha and beta ranking monkeys. Although we have not finished collecting data this summer, I performed preliminary analyses on data collected thus far using SPSS V.25 (IBM).
Preliminary Results
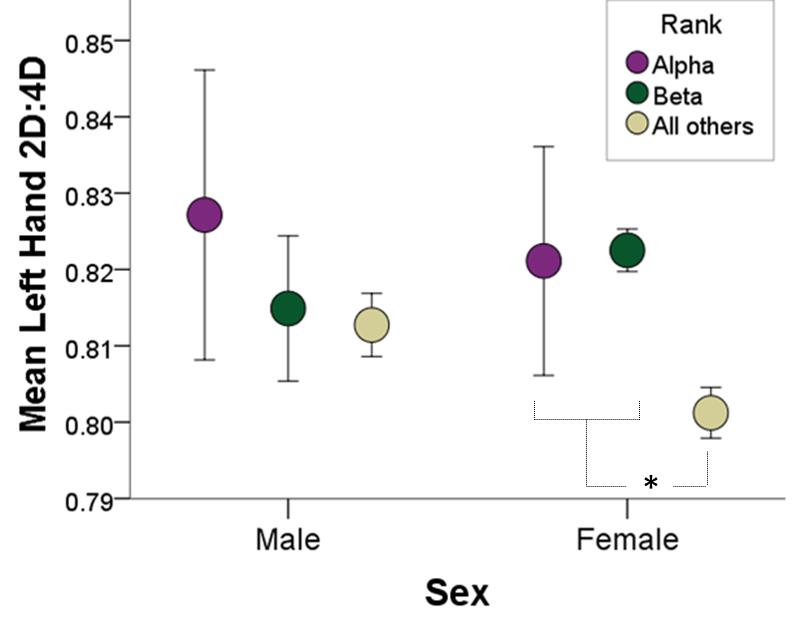
Figure 1: Alpha and Beta Females Masculinized Left-hand 2D:4D Ratio.
Discussion
As I hypothesized, based on the data I have collected thus far, I found that the alpha- and beta-ranking females had higher (i.e., for a rhesus monkey, more male-typical) left-hand 2D:4D ratio than the other females that were not alpha- or beta-ranking. One potential explanation for this finding is that the alpha- and beta-ranking females experienced higher levels of PAE than the other females did, potentially making these females more competitive or aggressive. Through these effects on personality and temperament, PAE may have driven these females to compete for and strive to maintain higher social dominance rank than they otherwise would have with lower levels of PAE. Although this interpretation is speculative, we plan to conduct follow up observations on these subjects to explore how alpha- and beta-ranking females behave, and whether levels of PAE really made them “destined to dominate.”
References
Abbott, A. D., Colman, R. J., Tiefenthaler, R., Dumesic, D. A., & Abbott, D. H. (2012). Early-to-mid gestation fetal testosterone increases right hand 2D∶4D finger length ratio in polycystic ovary syndrome-like monkeys. PLoS One, 7(8), e42372.
Baxter, A., Wood, E. K., Jarman, P. A., Cameron, A. N., Capitanio, J. P., & Higley, J. D. Sex Differences in rhesus monkeys’ digit ratio (2D:4D ratio) and its association with maternal social dominance rank. Frontiers in Neuroscience. In Press.
Manning, J. T. (2011). Resolving the role of prenatal sex steroids in the development of digit ratio. Proceedings of the National Academy of Sciences, 108(39), 16143-16144.
Nelson, E., Hoffman, C. L., Gerald, M. S., & Shultz, S. (2010). Digit ratio (2D:4D) and dominance rank in female rhesus macaques (Macaca mulatta). Behavioral Ecology and Sociobiology, 64(6), 1001-1009.
Zheng, Z., & Cohn, M. J. (2011). Developmental basis of sexually dimorphic digit ratios. Proceedings of the National Academy of Sciences, 108(39), 16289-16294.