
Storm Atwood and Dr. C. Brock Kirwan, Psychology
The prevalence of autism, a neurodevelopmental disorder impairing social interaction, communication, and behavior, has risen from 1 in 110 affected to 1 in 88 affected (Centers for Disease Control, 2012). The rise in prevalence of autism has increased the awareness of autism and increased the desire and need for more research. In general, it has been found that memory performance remains largely unchanged in autism. For example, a recent study on recognition memory in autism found that there was no difference in the ability of an autistic participant versus a control participant in identifying whether a stimulus was old or new (Massand et al., 2013). A similar study also found that children with autism appear to have enhanced visual recognition when viewing inanimate objects (Kuusikko-Gauffin et al., 2011). However, it is an open question why object recognition memory in autism is equal to or superior to non-autistic people. It has been suggested that memory processing may be more specific in autism (Sahay et al., 2011). This project seeks to further characterize memory processes in autism in order to determine whether memory representations are more specific as predicted by theorists. This project will increase the understanding of the quality of memory in autism and provide additional information to the body of research involving autism and memory. Another positive outcome of this study is that by increasing understanding of memory processes in autism, occupational and social therapy can be improved to better benefit those diagnosed with autism.
Seventy participants (25 diagnosed with Autism Spectrum Disorder, and 45 without any diagnosis) ages 17 to 36 were recruited for the study. These participants completed a pattern separation recognition memory task before being scanned using magnetic resonance imaging (MRI) at the BYU MRI Research Facility. In the task, participants were shown a series of objects, one at a time. After about 10 items the objects began to repeat. Participants selected whether each stimulus was “old” (previously seen or repeated), “similar” (lures similar to, but not exactly the same as a prior stimulus), or “new” (novel foils not previously encountered in the experiment). There were approximately 600 images viewed over blocks lasting 10 minutes; each image was shown for 2.5 seconds. Following the continuous recognition memory task, each participant contributed a standard-resolution structural MRI scan, a high resolution MRI scan focusing on the medial temporal lobe, and a diffusion tensor imaging (DTI) scan. These scans will allowed us to investigate the relationship between performance on the pattern separation task and 1) volumes of different brain structures, 2) white matter integrity, and 3) structural connectivity between the hippocampus and other medial temporal lobe structures.
The overall pattern separation (PS) score (calculated as the proportion of similar lures to which the participant responded “similar” corrected for the baseline “similar” response rate) did not differ between groups, t(66) = 0.66, p = .54. Figure 1 show that the autism spectrum disorder (ASD) group’s performance, Figure 2 shows the control group’s performance. There were no significant differences between the groups in their responses to novel foils. However, in the repeated and similar lures categories, the ASD group choose both “new” and “similar” significantly more often than controls, for “new” choices t(66) = 2.10, p = .04; for “similar” choices t(66) = 2.31, p = .02. The ASD group also chose “new” for the “similar” lure stimuli significantly more frequently than controls, t(66) = 2.29, p = .03. Analysis of the MRI data is ongoing.
The goal of this project was to explore whether memory discrimination, the ability to distinguish between similar stimuli, in autism is overactive or impaired. Overactive would indicate that memory is hyper-specific and that autistic participants would be able to correctly identify the differences between similar stimuli. Impaired would indicate that memory is not specific enough and the participants would be unable to correctly differentiate between similar stimuli. We did not observe a difference between ASD and control groups in their responses to similar lure stimuli, indicating that pattern separation processes are intact in ASD. However, we found that participants with autism were more likely to choose “new” and “similar” than the controls. This result was unexpected, but may be due to an overall response bias in ASD meant to avoid mistakes such as false alarming to a lure stimulus.
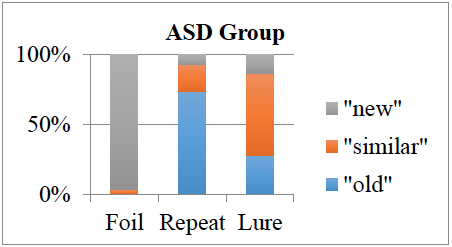
Figure 1: Choices for each stimulus category across diagnostic groups. Horizontal axis represents the type of stimulus shown on the monitor. Each section within the vertical columns represents the number of choices for each type chosen by the participant.
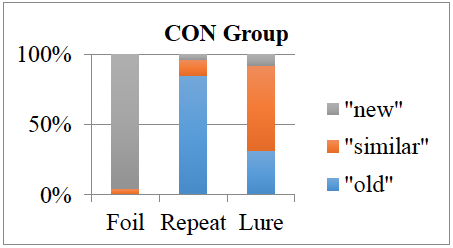
Figure 2: Choices for each stimulus category across diagnostic groups. Horizontal axis represents the type of stimulus shown on the monitor. Each section within the vertical columns represents the number of choices for each type chosen by the participant.
References
- Bakker, A., Kirwan, C.B., Miller, M.I., & Stark, C.E.L. (2008). Pattern Separation in the human hippocampal CA3 and dentate gyrus subfields. Science. 319(5870):1640-1642.
- Centers for Disease Control and Prevention. (2012). Prevalence of autism spectrum disorders–autism and developmental disabilities monitoring network, 14 sites, united states, 2008. Morbidity and Mortality Weekly Report, 61(3), 1-19.
- Kuusikko-Gauffin, S., Eira, J., Alice, C., Rachel, P., Katja, J., Marja-Leena, M., . . . Irma, M. (2011). Face memory and object recognition in children with high-functioning autism or asperger syndrome and in their parents. Research in Autism Spectrum Disorders, 5(1), 622-628.
- Massand, E., Bowler, D. M., Mottron, L., Hosein, A., & Jemel, B. (2013). Erp correlates of recognition memory in autism spectrum disorder. Journal of Autism & Childhood Schizophrenia, 43(9), 2038-2047.
- Sahay, A., Wilson, D., & Hen, R. (2011). Pattern separation: A common function for new neurons in hippocampus and olfactory bulb. Neuron, 70(4), 582-588.