Melanie A. Watson and Dr. Edwin Lephart, Zoology
Calbindin-D28K is a calcium-binding protein that serves a potentially important role in both the development and function of the central nervous system (CNS). Previous studies indicate the dimorphic expressions of calbindin within the medial basal hypothalamus (N4BH), supporting its putative importance in the neurogenesis of sexually dimorphic brain structures.2 Another primary action of this protein is to buffer intracellular calcium levels, thus functioning as a neuroprotective mechanism against pre-mature apoptosis. Regulation of calbindin is reported to be, in part, stimulated by glucocorticoids in response to activation of the hypothalamic-pituitaryadrenal stress axis.3 Evidence also exists for the potential regulation of calcium-binding proteins by androgens. In order to examine the influence of androgens upon hypothalamic regulation of calbindin-D28K, pregnant rats were treated with either testosterone or flutamide, a known androgen receptor blocker.
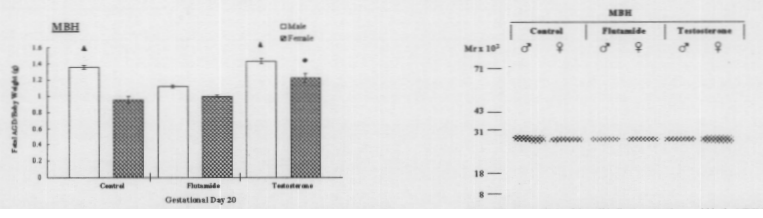
Pregnant rats were assigned to 3 treatment groups: 1) control (n=2), 2) flutanmide (n=3) and 3) testosterone (n=3). Injections (subcutaneous) were initiated on gestational day (GD) 17 with oil (testosterone) or ethanol (flutamide) as the injection vehicles and continued until GD20. Then the animals were killed on the afternoon of GD20. Thereafter, fetuses were removed from the uterine horns, and anogenital distance (AGD), body weight, and sex were determined. The effectiveness of the treatments was measured by analysis of fetal body weight as a function of AGD. The MBH region from 5 male and 5 female fetuses was collected from each litter. Brain samples were quantified for total protein content by the Lowery protein assay.4 Calbindin levels within the MIBH tissue samples were determined by Western Blot analysis. The immunoreactive bands were quantified by measuring optical density. The data were then tested by analysis of variance and compared against each other by pairwise comparison (via Tukey’s analysis) to determine significant differences across treatment groups as well as between male and female samples within each treatment.
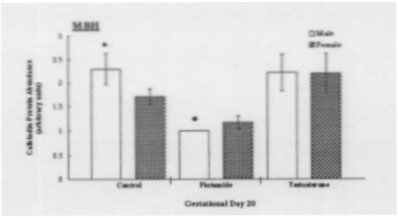
Following statistical analysis of the data, the following results were obtained. First, the treatments were effective (see Fig. 1). Testosterone is, in part, responsible for development of external sex characteristics, namely in this study, AGD. Accordingly, males have a larger AGD if compared to females. Male fetuses treated with flutamide exhibit significantly decreased AGD values compared to control values, where females were not significantly affected. In testosterone-treated animals, male AGD values were slightly increased above control values, whereas female AGD values were increased, thereby appearing more like those of male controls. Second, Western analysis for calbindin recognized a protein band with a molecular weight of 28,000 Daltons (see Fig. 2). Control males exhibit a higher abundance than control females, this being consistent with data previously reported.1 Flutamide male and female protein levels were decreased as compared to control male and female MBH samples, male protein levels were more dramatically affected. Compared to control samples, calbindin levels under the influence of testosterone increased in female samples and decreased slightly in male samples. Both male and female samples were significantly greater than flutamide male and female calbindin levels (see Fig. 3).
Based on the results of this study, that blocking the androgen supply with Flutamide dramatically decreases the amount of calbindin produced in male and female rat fetuses at GD20, calbindin appears to be regulated during prenatal development, in part, by an androgen-dependent mechanism. A possible explanation for the non-significant effect on calbindin production by testosterone treatments is supported by previous findings that report that while androgens are important for the development of sexually dimorphic brain structures, excess testosterone does not notably increase the volume of such structures. Accordingly, because calbindin, along with androgens, is thought to be involved in the development of sexually dimorphic structures, the possibility exists that while a certain amount of androgens may be required to maintain calbindin production, an excess amount may have no effect.
References
- Lephart, E.D. 1996. Developmental Brain Research 96:281-284.
- ILephart, E.D., M.A. Watson, N.A. Jacobson, et al. 1997. Developmental Brain Research 100:117-120.
- Lowry, O.H., N.J. Rosenbourgh, AL. Farr, et al. 1951. Journal of Biology and Chemistry 193:265-275.
- The aid of Heather Taylor, David Ladle, Nathan Jacobson, and Lori Mathias is gratefully acknowledged.